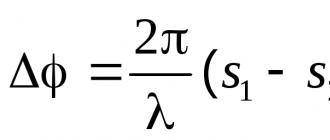Что такое мембранный потенциал. Мембранный потенциал действия
»: Потенциал покоя - это важное явление в жизни всех клеток организма, и важно знать, как он формируется. Однако это сложный динамический процесс, трудный для восприятия целиком, особенно для студентов младших курсов (биологических, медицинских и психологических специальностей) и неподготовленных читателей. Впрочем, при рассмотрении по пунктам, вполне возможно понять его основные детали и этапы. В работе вводится понятие потенциала покоя и выделяются основные этапы его формирования с использованием образных метафор, помогающих понять и запомнить молекулярные механизмы формирования потенциала покоя.
Мембранные транспортные структуры - натрий-калиевые насосы - создают предпосылки для возникновения потенциала покоя. Предпосылки эти - разность в концентрации ионов на внутренней и наружной сторонах клеточной мембраны. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка ионов калия (K +) выровнять свою концентрацию по обе стороны мембраны приводит к его утечке из клетки и потере вместе с ними положительных электрических зарядов, за счёт чего значительно усиливается общий отрицательный заряд внутренней поверхности клетки. Эта «калиевая» отрицательность составляет бóльшую часть потенциала покоя (−60 мВ в среднем), а меньшую его часть (−10 мВ) составляет «обменная» отрицательность, вызванная электрогенностью самого ионного насоса-обменника.
Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
Вы знаете, что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку» и почему она моментально не разряжается?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка - нейрон. Главное, что лежит в основе работы нейрона - это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы - это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя .
Определение понятия «потенциал покоя»
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал - мембранный потенциал покоя .
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии .
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e ), полный электрический заряд любого тела можно представить как q = ±N×e , где N - целое число.
Потенциал покоя - это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами - химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде - это ионные токи .
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи - положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K +), а внутри - отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K +).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они - отрицательные.
Термин «отрицательность», который мы будем применять для характеристики электрического потенциала внутри клетки, пригодится нам для простоты объяснения изменений уровня потенциала покоя. В этом термине ценно то, что интуитивно понятно следующее: чем больше отрицательность внутри клетки - тем ниже в отрицательную сторону от нуля смещён потенциал, а чем меньше отрицательность - тем ближе отрицательный потенциал к нулю. Это намного проще понять, чем каждый раз разбираться в том, что же именно означает выражение «потенциал возрастает» - возрастание по абсолютному значению (или «по модулю») будет означать смещение потенциала покоя вниз от нуля, а просто «возрастание» - смещение потенциала вверх к нулю. Термин «отрицательность» не создаёт подобных проблем неоднозначности понимания.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Так куда же деваются из клетки положительно заряженные частицы? Напомню, что это покинувшие клетку и скопившиеся снаружи ионы натрия - Na + - и калия - K + .
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
- вначале она обменивает «свой» натрий на «чужой» калий (да-да, одни положительные ионы на другие, такие же положительные);
- потом из неё происходит утечка этих «наменянных» положительных ионов калия, вместе с которыми из клетки утекают положительные заряды.
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
В мембране нервной клетки постоянно работают белковые насосы-обменники (аденозинтрифосфатазы, или Na + /K + -АТФазы), встроенные в мембрану. Они меняют «собственный» натрий клетки на наружный «чужой» калий.
Но ведь при обмене одного положительного заряда (Na +) на другой такой же положительный заряд (K +) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 - горький, 0,05–0,1 - горько-солёный, а начиная с 0,2 и выше - сложный вкус, состоящий из солёного, горького и кислого .
Важно здесь то, что обмен натрия на калий - неравный . За каждые отданные клеткой три иона натрия она получает всего два иона калия . Это приводит к потере одного положительного заряда при каждом акте ионного обмена. Так что уже на этом этапе за счёт неравноценного обмена клетка теряет больше «плюсов», чем получает взамен. В электрическом выражении это составляет примерно −10 мВ отрицательности внутри клетки. (Но помните, что нам надо ещё найти объяснение для оставшихся −60 мВ!)
Чтобы легче было запомнить работу насосов-обменников, образно можно выразиться так: «Клетка любит калий!» Поэтому клетка и затаскивает калий к себе, несмотря на то, что его и так в ней полно. И поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. И поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. (Вот что делает любовь, пусть она даже и не настоящая!)
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной - поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток .
Образно говоря, можно выразиться так: Создавая потенциал покоя, клетка «заряжается любовью». Это любовь к двум вещам:
- любовь клетки к калию (поэтому клетка насильно затаскивает его к себе);
- любовь калия к свободе (поэтому калий покидает захватившую его клетку).
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
- Дефицит натрия (Na +) в клетке.
- Избыток калия (K +) в клетке.
- Появление на мембране слабого электрического потенциала (−10 мВ).
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь : растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой - потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки . И это у него получается!
Ионы калия K + покидают клетку под действием химического градиента их концентрации по разные стороны мембраны (мембрана значительно более проницаема для K + , чем для Na +) и уносят с собой положительные заряды. Из-за этого внутри клетки нарастает отрицательность.
Тут ещё важно понять то, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов надо по отдельности рассматривать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно - концентрацию по калию внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это, бывает, делается в учебниках.
По закону выравнивания химических концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку; туда же его влечёт и электрическая сила (как мы помним, цитоплазма заряжена отрицательно). Хотеть-то он хочет, но не может, так как мембрана в обычном состоянии плохо его пропускает. Натриевые ионные каналы, имеющиеся в мембране, в норме закрыты. Если все же его заходит немножко, то клетка сразу же обменивает его на наружный калий с помощью своих натрий-калиевых насосов-обменников. Получается, что ионы натрия проходят через клетку как бы транзитом и не задерживаются в ней. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Он выходит наружу через особые каналы в мембране - «калиевые каналы утечки», которые в норме открыты и выпускают калий .
К + -каналы утечки постоянно открыты при нормальных значениях мембранного потенциала покоя и проявляют взрывы активности при сдвигах мембранного потенциала, которые длятся несколько минут и наблюдаются при всех значениях потенциала. Усиление К + -токов утечки ведёт к гиперполяризации мембраны, тогда как их подавление - к деполяризации. ...Однако, существование канального механизма, ответственного за токи утечки, долгое время оставалось под вопросом. Только сейчас стало ясно, что калиевая утечка - это ток через специальные калиевые каналы.
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) .
От химического - к электрическому
А теперь - ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов .
Калий (K +) положительно заряжен, и поэтому он, когда выходит из клетки, выносит из неё не только самого себя, но и положительный заряд. За ним изнутри клетки к мембране тянутся «минусы» - отрицательные заряды. Но они не могут просочиться через мембрану - в отличие от ионов калия - т.к. для них нет подходящих ионных каналов, и мембрана их не пропускает. Помните про оставшиеся необъяснёнными нами −60 мВ отрицательности? Это и есть та самая часть мембранного потенциала покоя, которую создаёт утечка ионов калия из клетки! И это - большая часть потенциала покоя.
Для этой составной части потенциала покоя есть даже специальное название - концентрационный потенциал . Концентрационный потенциал - это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия .
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста , по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал E K:

где Е к - равновесный потенциал, R - газовая постоянная, Т - абсолютная температура, F - постоянная Фарадея, К + внеш и K + внутр - концентрации ионов К + снаружи и внутри клетки, соответственно. По формуле видно, что для расчёта потенциала между собой сравниваются концентрации ионов одного вида - K + .
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей Р K: PNa :P Cl = 1: 0,04: 0,45 .
Заключение
Итак, поте нциал покоя состоит из двух частей:
- −10 мВ , которые получаются от «несимметричной» работы мембранного насоса-обменника (ведь он больше выкачивает из клетки положительных зарядов (Na +), чем закачивает обратно с калием).
- Вторая часть - это всё время утекающий из клетки калий, уносящий положительные заряды. Его вклад - основной: −60 мВ . В сумме это и дает искомые −70 мВ.
Что интересно, калий перестанет выходить из клетки (точнее, его вход и выход уравниваются) только при уровне отрицательности клетки −90 мВ. В этом случае сравняются химические и электрические силы, проталкивающие калий через мембрану, но направляющие его в противоположные стороны. Но этому мешает постоянно подтекающий в клетку натрий, который несёт с собой положительные заряды и уменьшает отрицательность, за которую «борется» калий. И в итоге в клетке поддерживается равновесное состояние на уровне −70 мВ.
Вот теперь мембранный потенциал покоя окончательно сформирован.
Схема работы Na + /K + -АТФазы наглядно иллюстрирует «несимметричный» обмен Na + на K + : выкачивание избыточного «плюса» в каждом цикле работы фермента приводит к отрицательному заряжению внутренней поверхности мембраны. Чего в этом ролике не сказано, так это того, что АТФаза ответственна за менее чем 20% потенциала покоя (−10 мВ): оставшаяся «отрицательность» (−60 мВ) появляется за счет выхода из клетки через «калиевые каналы утечки» ионов K + , стремящихся выровнять свою концентрацию внутри клетки и вне нее.
Литература
- Jacqueline Fischer-Lougheed, Jian-Hui Liu, Estelle Espinos, David Mordasini, Charles R. Bader, et. al.. (2001). Human Myoblast Fusion Requires Expression of Functional Inward Rectifier Kir2.1 Channels . J Cell Biol . 153 , 677-686;
- Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. (1998). Role of an inward rectifier K + current and of hyperpolarization in human myoblast fusion . J. Physiol. 510 , 467–476;
- Sarah Sundelacruz, Michael Levin, David L. Kaplan. (2008). Membrane Potential Controls Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells . PLoS ONE . 3 , e3737;
- Павловская М.В. и Мамыкин А.И. Электростатика. Диэлектрики и проводники в электрическом поле. Постоянный ток / Электронное пособие по общему курсу физики. СПб: Санкт-Петербургский государственный электротехнический университет;
- Ноздрачёв А.Д., Баженов Ю.И., Баранникова И.А., Батуев А.С. и др. Начала физиологии: Учебник для вузов / Под ред. акад. А.Д. Ноздрачёва. СПб: Лань, 2001. - 1088 с.;
- Макаров А.М. и Лунева Л.А. Основы электромагнетизма / Физика в техническом университете. Т. 3;
- Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология). Казань: Арт-кафе, 2010. - 271 с.;
- Родина Т.Г. Сенсорный анализ продовольственных товаров. Учебник для студентов вузов. М.: Академия, 2004. - 208 с.;
- Кольман Я. и Рем К.-Г. Наглядная биохимия. М.: Мир, 2004. - 469 с.;
- Шульговский В.В. Основы нейрофизиологии: Учебное пособие для студентов вузов. М.: Аспект Пресс, 2000. - 277 с..
А. Характеристика ПД. ПД - электрический процесс, выражающийся в быстром колебании мембранного потенциала вследствие перемещения ионов в клетку и т клетки и способный распространяться без затухания (без декремента). Он обеспечивает передачу сигналов между нервными клетками, между нервными центрами и рабочими органами, в мышцах - процесс электромеханического сопряжения (рис. 3.3, а).
Величина ПД нейрона колеблется в пределах 80-110 мВ, длительность пика ПД нервного волокна составляет 0,5-1 мс. Амплитуда ПД не зависит от силы раздражения, она всегда максимальна для данной клетки в конкретных условиях: ПД подчиняется закону «все или ничего», но не подчиняется закону силовых отношений -закону силы. ПД либо совсем не возникает на раздражение клетки, если оно мало, либо он максимальной величины, если раздражение является пороговым или сверхпороговым. Следует отметить, что слабое (подпороговое) раздражение может вызвать локальный потенциал. Он подчиняется закону силы: с увеличением силы стимула величина его возрастает (подробнее см. раздел 3.6). В составе ПД различают три фазы: 1 фаза - деполяризация, т.е. исчезновение заряда клетки - уменьшение мембранного потенциала до нуля; 2 фаза - инверсия, изменение заряда клетки на обратный, когда внутренняя сторона мембраны клетки заряжается положительно, а внешняя - отрицательно (от лат. туегзю - переворачивание); 3 фаза - реполяризация, восстановление исходного заряда клетки, когда внутренняя поверхность клеточной мембраны снова заряжается отрицательно, а наружная - положительно.
Б. Механизм возникновения ПД. Если действие раздражителя на клеточную мембрану приводит к возникновению ПД, далее сам процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение иона Ка + в клетку, а иона К + - из клетки. Величина мембранного потенциала при этом сначала уменьшается, а затем снова восстанавливается до исходного уровня. На экране осциллографа отмеченные изменения мембранного потенциала предстают в виде пикового потенциала - ПД. Он возникает вследствие накопленных и поддерживаемых ионными насосами градиентов концентраций ионов внутри и вне клетки, т.е. за счет потенциальной энергии в виде электрохимических градиентов разных ионов. Если заблокировать процесс выработки энергии, то ПД некоторый период времени будут возникать, но после исчезновения градиентов концентраций ионов (устранение потенциальной энергии) клетка генерировать ПД не будет. Рассмотрим фазы ПД.
| Рис. 3.3. Схема, отражающая процесс возбуждения. а - потенциал действия, его фазы: 1 - деполяризация, 2 - инверсия (овершут), 3 - реполяризация, 4 - следовая гиперполяризация; б - натриевые ворота; (Ь-1 - в состоянии покоя клетки); в - калиевые ворота (1 - в состоянии покоя клетки). Знаки плюс (+) и минус (-) - знаки заряда внутри и вне клетки в различные фазы ПД. (См. пояснения в тексте.) Существует много различных названий фаз ПД (единого мнения не сложилось): 1) местное возбуждение - пик ПД - следовые потенциалы; 2) фаза нарастания - фаза спада -следовые потенциалы; 3) деполяризация - овершут (перехлест, превышение, перелет), причем эта фаза в свою очередь делится на две част: восходящая (инверсия, ОТ лат. шуегяю - переворачивание) н нисходящая (реверсия, от лат. геуегзю - возврат) - реполя-рнзапия. Имеются и другие названия. |
Отметим одно противоречие: термины «реполяризация» и «реверсия» но смыслу одинаковы - возврат к предыдущему состоянию, но эти состояния различны: в одном случае заряд исчезает (реверсия), в другом -восстанавливается (реполяршация). Наиболее корректны тс названия фаз ПД, в которых заложена общая идея, например изменение заряда клетки. В этой связи обоснованно использовать следующие названия фаз ПД: !) фаза деполяризации - процесс исчезновения заряда клетки до нуля; 2) фаза инверсии - изменение заряда клетки на противоположный. т. е. весь период ПД, когда внутри клетки заряд положительный, а снаружи - отрицательный; 3) фаза реполярпзацин - восстановление заряда клетки до исходной величины (возврат к потенциалу покоя).
1. Фаза деполяризации (см. рис. 3.3, а, 1). При действии деполяризующего раздражителя на клетку (медиатор, электрический ток) вначале уменьшение мембранного потенциала (частичная деполяризация) происходит без изменения проницаемости мембраны для ионов. Когда деполяризация достигает примерно 50% пороговой величины (порогового потенциала), возрастает проницаемость ее мембраны для иона Ка + , причем в первый момент сравнительно медленно. Естественно, что скорость входа ионов Ка* в клетку при этом невелика. В этот период, как и во время всей фазы деполяризации, движущей силой, обеспечивающей вход иона Na + в клетку, являются концентрационный и электрический градиенты. Напомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация ионов Na+ вне клетки в 10-12 раз больше, чем внутри клетки. При возбуждении нейрона повышается проницаемость его мембраны и для ионов Са+, но его ток в клетку значительно меньше, чем ионов Nа + . Условием, обеспечивающим вход иона Nа + в клетку и последующий выход иона К* из клетки, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма ионных Nа- и К-каналов. Длительность пребывания электроуправляемого канала в открытом состоянии носит вероятностный характер и зависит от величины мембранного потенциала. Суммарный ток ионов в любой момент определяется числом открытых каналов клеточной мембраны. Воротный механизм ^-каналов расположен на внешней стороне клеточной мембраны (Na+ движется внутрь клетки), воротный механизм К-каналов -на внутренней (К + движется из клетки наружу).
Активация Nа- и К-каналов (открытие ворот) обеспечивается уменьшением мембранного потенциала, Когда деполяризация клетки достигает критической величины (E kp , критический уровень деполяризации - КУД), которая обычно составляет -50 мВ (возможны и другие величины), проницаемость мембраны для ионов Nа + резко возрастает - открывается большое число по-тенциалзависимых ворот Nа-каналов и ионы Nа + лавиной устремляются в клетку. В результате интенсивного тока ионов Nа + внутрь клетки далее процесс деполяризации проходит очень быстро. Развивающаяся деполяризация клеточной мембраны вызывает дополнительное увеличение ее проницаемости и, естественно, проводимости ионов Na+ - открываются все новые и новые активационные т-ворота Nа-каналов, что придает току ионов Na* в клетку характер регенеративного процесса. В итоге ПП исчезает, становится равным нулю. Фаза деполяризации на этом заканчивается.
2. Фаза инверсии. После исчезновения ПП вход Nа+ в клетку продолжается (m - ворота Na-каналов еще открыты - h-2), поэтому число положительных ионов в клетке превосходит число отрицательных, заряд внутри клетки становится положительным, снаружи - отрицательным. Процесс перезарядки мембраны представляет собой 2-ю фазу ПД - фазу инверсии (см. рис. 3.3, в, 2). Теперь электрический градиент препятствует входу Na+ внутрь клетки (положительные заряды отталкиваются друг от друга), проводимость Na* снижается. Тем не менее некоторый период (доли миллисекунды) ионы Na + продолжают входить в клетку, об этом свидетельствует продолжающееся нарастание ПД. Это означает, что концентрационный градиент, обеспечивающий движение ионов Ка + в клетку, сильнее электрического, препятствующего входу ионов Nа* в клетку. Во время деполяризации мембраны увеличивается проницаемость ее и для ионов Са 2+ , они также идут в клетку, но в нервных клетках роль ионов Са 2+ в развитии ПД мала. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом ионов Nа* в клетку.
Примерно через 0,5-1 мс после начала деполяризации рост ПД прекращается вследствие закрытия ворот Ка-каналов (Ь-3) и открытия ворот К-каналов (в, 2), т.е. увеличения проницаемости для ионов К + . Поскольку ионы К + находятся преимущественно внутри клетки, они, согласно концентрационному градиенту, быстро выходят из клетки, вследствие чего в клетке уменьшается число положительно заряженных ионов. Заряд клетки начинает возвращаться к исходному уровню. В фазу инверсии выходу ионов К* из клетки способствует также электрический градиент. Ионы К* выталкиваются положительным зарядом из клетки ипритягиваются отрицательным зарядом снаружи клетки. Так продолжается до полного исчезновения положительного заряда внутри клетки - до конца фазы инверсии (см. рис. 3.3, а - пунктирная линия), когда начинается следующая фаза ПД - фаза реполяризации. Калий выходит из клетки не только по управляемым каналам, ворота которых открыты, но и по неуправляемым каналам утечки.
Амплитуда ПД складывается из величины ПП (мембранный потенциал покоящейся клетки) и величины фазы инверсии - около 20 мв. Если мембранный потенциал в состоянии покоя клетки мал, то амплитуда ПД этой клетки будет небольшой.
3. Фаза реполяризации. В этой фазе проницаемость клеточной мембраны для ионов К + все еще высока, ионы К + продолжают быстро выходить из клетки согласно концентрационному градиенту. Клетка снова внутри имеет отрицательный заряд, а снаружи - положительный (см. рис. 3.3, а, 3), поэтому электрический градиент препятствует выходу К* из клетки, что снижает его проводимость, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее действия электрического градиента. Таким образом, вся нисходящая часть пика ПД обусловлена выходом иона К + из клетки. Нередко в конце ПД наблюдается замедление реполяризации, что объясняется уменьшением проницаемости клеточной мембраны для ионов К + и замедлением выхода их из клетки вследствие закрытия ворот К-каналов. Другая причина замедления тока ионов К + связана с возрастанием положительного потенциала наружной поверхности клетки и формированием противоположно направленного электрического градиента.
Главную роль в возникновении ПД играет ион Na*, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. При замене иона Nа + в среде на другой ион, например холин, или в случае блокировки Na-каналов тетродотоксином, ПД в нервной клетке не возникает. Однако проницаемость мембраны для иона К + тоже играет важную роль. Если повышение проницаемости для иона К + предотвратить тетраэтиламмонием, то мембрана после ее деполяризации реполяризуется гораздо медленнее, только за счет медленных неуправляемых каналов (каналы утечки ионов), через которые К + будет выходить из клетки.
Роль ионов Са 2+ в возникновении ПД в нервных клетках незначительна, в некоторых нейронах она существенна, например в дендритах клеток Пуркинье мозжечка.
В. Следовые явления в процессе возбуждения клетки. Эти явления выражаются в гиперполяризации или частичной деполяризации клетки после возвращения мембранного потенциала к исходной величине (рис. 3.4).
Следовая гиперполяризация клеточной мембраны обычно является следствием еще сохраняющейся повышенной проницаемости клеточной мембраны для К + . Ворота К-каналов еще не полностью закрыты, поэтому К + продолжает выходить из клетки согласно концентрационному градиенту, что и ведет к гиперполяризации клеточной мембраны. Постепенно проницаемость клеточной мембраны возвращается к исходной (натриевые и калиевые ворота возвращаются в исходное состояние), а мембранный потенциал становится таким же, каким он был до возбуждения клетки. Ионные помпы непосредственно за фазы потенциала действия не отвечают, ионы перемещаются с огромной скоростью согласно концентрационному и частично электрическому градиентам.
Следовая деполяризация также характерна для нейронов. Механизм ее изучен недостаточно. Возможно, она обусловлена кратковременным повышением проницаемости клеточной мембраны для Ка* и входом его в клетку согласно концентрационному и электрическому градиентам.
Наиболее растпространенный метод изучения функций ионных каналов - метод фиксации напряжения (voltage-clamp). Мембранный потенциал с помощью подачи электрического напряжения изменяют и фиксируют на определенном уровне, затем клеточную мембрану градуально деполяризуют, что ведет к открытию ионных каналов и возникновению ионного тока, который мог бы деполяризовать клетку. При этом пропускают электрический ток, равный по величине, но противоположный по знаку ионному току, поэтому трансмембранная разность потенциалов не изменяется. Это позволяет изучить величину ионного тока через мембрану. Применение различных блокаторов ионных каналов дает дополнительную возможность более глубоко изучить свойства каналов.
Количественное соотношение между ионными токами по отдельным каналам в покоящейся клетке и во время ПД и их кинетику можно выяснить с помощью метода локальной фиксации потенциала (patch-clamp). К мембране подводят микроэлектрод - присоску (внутри его создается разрежение) и, если на этом участке оказывается канал, исследуют ионный ток через него. В остальном методика подобна предыдущей. И в этом случае применяют специфические блокаторы каналов. В частности, при подаче на мембрану фиксированного деполяризующего потенциала было установлено, что через Ка-каналы может проходить и ион К + , но его ток в 10-12 раз меньше, а через К-каналы может проходить ион Ма + , его ток в 100 раз меньше, чем ток ионов К + .
Запас ионов в клетке, обеспечивающий возникновение возбуждения (ПД), огромен. Концентрационные градиенты ионов в результате одного цикла возбуждения практически не изменяются. Клетка может возбуждаться до 5 * 10 5 раз без подзарядки, т.е. без работы Ма/К-насоса. Число импульсов, которое генерирует и проводит нервное волокно, зависит от его толщины, что определяет запас ионов. Чем толще нервное волокно, тем больше запас ионов, тем больше импульсов оно может генерировать (от нескольких сотен до миллиона) без участия Nа/К-насоса. Однако в тонких волокнах на возникновение одного ПД расходуется около 1% концентрационных градиентов ионов Nа + и К*. Если заблокировать выработку энергии, то клетка будет еще многократно возбуждаться. В реальной действительности Nа/К-насос постоянно переносит ионы Nа + из клетки, а ионы К + возвращает в клетку, в результате чего поддерживается концентрационный градиент Nа + и К + за счет непосредственного расхода энергии, источником которой является АТФ. Имеются данные, что увеличение внутриклеточной концентрации Nа + сопровождается повышением интенсивности работы Nа/К-насоса. Это может быть связано исключительно с тем, что для переносчика становится доступно большее количество внутриклеточных ионов Na + .
В 1786 году профессор анатомии Болонского университета Луиджи Гальвани провел ряд опытов, положивших начало целенаправленным исследованиям в области биоэлектрических явлений. В первом опыте он подвешивал препарат обнаженных лапок лягушки с помощью медного крючка на железной решетке, и обнаружил, что при каждом касании мышцами решетки, они сокращались. Гальвани предположил, что сокращения мышц вообще – следствие воздействия на них «животного электричества», источником которого являются нервы и мышцы. Однако, по мнению Вольта, причиной сокращения был электрический ток, возникший в области контакта разнородных металлов. Гальвани поставил второй опыт, в котором источником тока, действовавшего на мышцу, точно был нерв: мышца опять сокращалась. Таким образом, было получено точное доказательство существования «животного электричества».
Все клетки имеют свой электрический заряд, который формируется в результате неодинаковой проницаемости мембраны для различных ионов. Клетки возбудимых тканей (нервная, мышечная, железистая) отличаются тем, что они под действием раздражителя меняют проницаемость своей мембраны для ионов, в результате чего ионы очень быстро транспортируются согласно электрохимическому градиенту. Это и есть процесс возбуждения. Его основой является потенциал покоя.
Потенциал покоя
Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах от -30 до -90 мВ. Внутренняя сторона мембраны в покое заряжена отрицательно, а наружная – положительно из-за неодинаковых концентраций катионов и анионов внутри и вне клетки.
Внутри- и внеклеточные концентрации ионов (ммоль/л) в мышечных клетках теплокровных животных
В нервных клетках похожая картина. Таким образом, видно, что основную роль в создании отрицательного заряда внутри клетки играют ионы K + и высокомолекулярные внутриклеточные анионы, главным образом они представлены белковыми молекулами с отрицательно заряженными аминокислотами (глутамат, аспартат) и органическими фосфатами. Эти анионы, как правило, не могут транспортироваться через мембрану, создавая постоянный отрицательный внутриклеточный заряд. Во всех точках клетки отрицательный заряд практически одинаков. Заряд внутри клетки является отрицательным как абсолютно (в цитоплазме анионов больше, чем катионов), так и относительно наружной поверхности клеточной мембраны. Абсолютная разность невелика, однако этого достаточно для создания электрического градиента.
Главным ионом, обеспечивающим формирование потенциала покоя (ПП), является K + . В покоящейся клетке устанавливается динамическое равновесие между числом входящих и выходящих ионовK + . Это равновесие устанавливается тогда, когда электрический градиент уравновесит концентрационный. Согласно концентрационному градиенту, создаваемому ионными насосами, K + стремится выйти из клетки, однако отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточной мембраны препятствуют этому (электрический градиент). В случае равновесия на клеточной мембране устанавливается равновесный калиевый потенциал.
Равновесный потенциал для каждого иона можно рассчитать по формуле Нернста:
E ion =RT/ZF·ln( o / i),
где E ion - потенциал, создаваемый данным ионом;
R – универсальная газовая постоянная;
Т – абсолютная температура (273+37°С);
Z – валентность иона;
F – постоянная Фарадея (9,65·10 4);
O – концентрация иона во внешней среде;
I - концентрация иона внутри клетки.
При температуре 37°С равновесный потенциал для K + равен -97мВ. Однако реальный ПП меньше – около -90 мВ. Это объясняется тем, что в формирование ПП свой вклад вносят и другие ионы. В целом ПП – это алгебраическая сумма равновесных потенциалов всех ионов, находящихся внутри и вне клетки, включающий также значения поверхностных зарядов самой клеточной мембраны.
Вклад Na + и Cl - в создание ПП невелик, но, тем не менее, он имеет место. В покое вход Na + в клетку низкий (намного ниже, чем K +), но он уменьшает мембранный потенциал. Влияние Cl - противоположно, так как это анион. Отрицательный внутриклеточный заряд не позволяет большому количеству Cl - проникнуть в клетку, поэтому Cl - это в основном внеклеточный анион. Как внутри клетки, так и вне ееNa + и Cl - нейтрализуют друг друга, вследствие чего их совместное поступление в клетку не оказывает существенного влияния на величину ПП.
Наружная и внутренняя стороны мембраны несут на себе собственные электрические заряды, преимущественно с отрицательным знаком. Это полярные составляющие мембранных молекул – гликолипидов, фосфолипидов, гликопротеинов. Ca 2+ , как внеклеточный катион, взаимодействует с наружными фиксированными отрицательными зарядами, а также с отрицательными карбоксильными группами интерстиция, нейтрализуя их, что приводит к увеличению и стабилизации ПП.
Для создания и поддержания электрохимических градиентов необходима постоянная работа ионных насосов. Ионный насос – это транспортная система, обеспечивающая перенос иона вопреки электрохимическому градиенту, с непосредственными затратами энергии. Градиенты Na + и K + поддерживаются с помощью Na/K – насоса. Сопряженность транспорта Na + и K + примерно в 2 раза уменьшает энергозатраты. В целом же траты энергии на активный транспорт огромны: лишь Na/K – насос потребляет около 1/3 всей энергии, расходуемой организмом в покое. 1АТФ обеспечивает один цикл работы – перенос 3Na + из клетки, и 2 K + в клетку. Асимметричный перенос ионов способствует заодно формированию и электрического градиента (примерно 5 – 10мВ).
Нормальная величина ПП является необходимым условием возникновения возбуждения клетки, т.е. распространения потенциала действия, инициирующего специфическую деятельность клетки.
Потенциал действия (ПД)
ПД – это электрофизиологический процесс, выражающийся в быстром колебании мембранного потенциала, вследствие специфического перемещения ионов и способный распространяться без декремента на большие расстояния. Амплитуда ПД колеблется в пределах 80 – 130 мВ, длительность пика ПД в нервном волокне – 0,5 – 1 мс. Амплитуда потенциала действия не зависит от силы раздражителя. ПД либо совсем не возникает, если раздражение подпороговое, либо достигает максимальной величины, если раздражение пороговое или сверхпороговое. Главным в возникновении ПД является быстрый транспорт Na + внутрь клетки, что способствует вначале снижению мембранного потенциала, а затем – изменению отрицательного заряда внутри клетки на положительный.
В составе ПД различают 3 фазы: деполяризацию, инверсию, и реполяризацию.
1. Фаза деполяризации . При действии на клетку деполяризующего раздражителя начальная частичная деполяризация происходит без изменения ее проницаемости для ионов (не происходит движение Na + внутрь клетки, т. к. закрыты быстрые потенциалчувствительные каналы для Na +). Na + - каналы обладают регулируемым воротным механизмом, который расположен на внутренней и внешней сторонах мембраны. Имеются активационные ворота (m – ворота) и инактивационные (h – ворота). В покое m – ворота закрыты, а h – ворота открыты. В мембране также имеются K + - каналы, имеющие только одни ворота (активационные), закрытые в покое.
Когда деполяризация клетки достигает критической величины (Е кр – критический уровень деполяризации, КУД), которая обычно равна 50мВ, проницаемость для Na + резко возрастает – открывается большое количество потенциалзависимых m – ворот Na + - каналов. За 1 мс через 1 открытый Na + - канал в клетку попадает до 6000 ионов. Развивающаяся деполяризация мембраны вызывает дополнительное увеличение ее проницаемости для Na + , открываются все новые и новые m - ворота Na + - каналы, так что ток Na + имеет характер регенеративного процесса (сам себя усиливает). Как только ПП становится равным нулю, фаза деполяризации заканчивается.
2.Фаза инверсии. Вход Na + в клетку продолжается, т. к. m - ворота Na + - каналы еще открыты, поэтому внутри клетки заряд становится положительным, а снаружи – отрицательным. Теперь электрический градиент препятствует входу Na + в клетку, однако, из-за того, что концентрационный градиент сильнее электрического, Na + все же проходит в клетку. В тот момент, когда ПД достигает максимального значения, происходит закрытие h – ворот Na + - каналов (эти ворота чувствительны к величине положительного заряда в клетке) и поступление Na + в клетку прекращается. Одновременно открываются ворота K + - каналов. K + транспортируется из клетки согласно химическому градиенту (на нисходящей фазе инверсии – еще и по электрическому градиенту). Выход положительных зарядов из клетки приводит к уменьшению ее заряда. K + с небольшой скоростью может выходить из клетки также через неуправляемые K + - каналы, которые всегда открыты. Все рассмотренные процессы являются регенеративными. Амплитуда ПД складывается из величины ПП и величины фазы инверсии. Фаза инверсии заканчивается, когда электрический потенциал снова становится равным нулю.
3.Фаза реполяризации. Связана с тем, что проницаемость мембраны для K + еще высока, и он выходит из клетки по градиенту концентрации, несмотря на противодействие электрического градиента (клетка внутри снова имеет отрицательный заряд). Выходом K + обусловлена вся нисходящая часть пика ПД. Нередко в конце ПД наблюдается замедление реполяризации, кто связано с закрытием значительной части ворот K + - каналов, а также – с возрастанием противоположно направленного электрического градиента.
Мембранный потенциал покоя (МПС) — это разность потенциалов между внешней и внутренней сторонами мембраны в условиях, когда клетка не возбуждено. Цитоплазма клетки заряжена отрицательно к внеклеточной жидкости неравномерным распределением анионов и катионов по обе стороны мембраны. Разность потенциалов (напряжение) для различных клеток имеет значение от -50 до -200 мВ (минус означает, что внутри клетка более негативно заряжена, чем снаружи). Мембранный потенциал покоя возникает на мембранах всех клеток — возбуждающих (нервов, мышц, секреторных клеток) и незбудливих.
МПС необходим для поддержания возбудимости таких клеток, как мышечные и нервовои. Также он влияет на транспорт всех заряженных частиц в любом типе клеток: он способствует пассивному транспорта анионов из клетки и катионов в клетку.
Образование и поддержания мембранного потенциала обеспечивают различные типы ионных насосов (в частности натрий-калиевый насос или натрий-калиевая АТФаза) и ионных каналов (калиевые, натриевые, хлорные ионные каналы).
Регистрация потенциала покоя
Для регистрации потенциала покоя используют специальную микроэлектродную технику. Микроэлектрод — это тоненькая стеклянная трубочка, с вытянутым концом, диаметром менее 1 мкм, заполненная раствором электролита (чаще хлорида калия). Рефернтним электродом служит серебряная хлорированная пластинка, расположенная в внеклеточном пространстве, оба электрода подключены к осциллографа. Сначала оба электрода занходяться в внеклеточном пространстве и разность потенциалов между ними отсутствует, если ввести регистрирующий микроэлектрод через мембрану в клетку, то осциллограф покажет скачкообразное смещение потенциала примерно до -80 мВ. Этот сдвиг потенциала называют мембранным потенциалом покоя.
Формирование потенциала покоя
К возникновению мембранного потенциала покоя приводят два фактора: во-первых, концентрации различных ионов отличаются внешне и всереднини клетки, во-вторых мембрана является полупроницаемой: одни ионы могут через нее проникать, другие — нет. Оба эти явления зависят от наличия в мембране специальных белков: концентрационные градиенты создают ионные насосы, а проницаемость мембраны для ионов обеспечивают ионные каналы. Важнейшую роль в формировании мембранного потенциала играют ионы калия, натрия и хлора. Концентрации этих ионов видризняюються по обе стороны мембраны. Для нейрона млекопитающих концентрация K + составляет 140 ммоль внутри клетки и только 5 мМ извне, градиент концентрации Na + почти противоположный — 150 ммоль снаружи и 15 мМ внутри. Такое распределение ионов поддерживается натрий-калиевым насосом в плазматической мембране — белком использующий энергию АТФ для закачки K + в клетку и скачивания Na + из нее. Также существует концентрационный градиент и для других ионов, например, хлорид аниона Cl -.
Концентрационные градиенты катионов калия и натрия — это химическая форма потенциальной энергии. В преобразовании энергии в электрическую участвуют ионные каналы — поры формируются скоплениями специальных трансмембранных белков. Когда ионы диффундируют через канал, они переносят единицу электрического заряда. Любой суммарный движение положительных или отрицательных ионов через мембрану будет создавать напряжение, или разность потенциалов по обе стороны мембраны.
Ионные каналы, участвующие в утовренни МПС имеют избирательную проницаемость, то есть дают возможность проникать только определенному типу ионов. В мембране нейрона в состоянии покоя открытые калиевые каналы (те, что в основном пропускают только калий), большинство натриевых каналов — закрыты. Диффузия ионов K + через калиевые каналы является решающим для создания мембранного потенциала. Так как концентрация K + значительно выше внутри клетки, химический градиент способствует оттоку этих катионов из клетки, поэтому в цитоплазме начинают преобладать анионы, которые не могут проходить через калиевые каналы.
Отток ионов калия из клетки ограничен самым мембранным потенциалом, поскольку при определенном его уровне накопление отрицательных зарядов в цитоплазме будет ограничивать движение катионов за пределы клетки. Таким образом, главным фактором в возникновении МПС является распределение ионов калия под действием электрического и химического потенциалов.
Равновесный потенциал
Для того, чтобы определить влияние движения определенного иона через полупроницаемую мембрану на формирование мембранного потенциала, строят модельные системы. Такая модельная система состоит из сосуда разделенной на две ячейки искусственной полупроницаемой мембраной, в которую встроены ионные каналы. В каждую ячейку можно погрузить электрод и померить разность потенциалов.
Рассмотрим случай, когда искусственная мембрана проницаема только для калия. По две стороны мембраны модельной системы создают градиент концентрации аналогичный таковому у нейроне: в ячейку, соответствующую цитоплазме (внутренняя ячейка), помещают 140 мМ раствор хлорида калия (KCl), в ячейку, соответствующую межклеточной жидкости (внешняя ячейка) — 5 ммоль раствор KCl. Ионы калия будут диффундировать через мембрану во внешнюю ячейку по градиенту концентрации. Но поскольку анионы Cl — проникать через мембрану не могут во внутренней ячейке возникать избыток отрицательного заряда, который будет препятствовать подали оттока катионов. Когда такие модельные нейроны достигнут состояния равновесия, действие химического и электрического потенциала будет сбалансирована, ни суммарной диффузии К + не будет наблюдаться. Значение мембранного потенциала, виинкае при таких условиях, называется равновесным потенциалом для определенного иона (Е ион). Равновесный потенциал для калия составляет примерно -90 мВ.
Аналогичный опыт можно провести и для натрия, установив между ячейками мембрану проникающей только для этого катиона, и поместив во внешнюю ячейку раствор хлорида натрия с концентрацией 150 мМ, а во внутреннюю — 15 мМ. Натрий будет двигаться во внутреннюю ячейку, ривоноважний потенциал для него составит примерно 62 мВ.
Количество ионов, должна диффундировать для генерации электрического потенциала очень невелика (примерно 10 -12 моль К + на 1 см 2 мембраны), этот факт имеет два важных последствия. Во-прешь, это означает, что концентрации ионов, которые могут проникать через мембрану, остаются стабильными снаружи и внутри клетки, даже после того как их движение обеспечил утоврення электрического потенциала. Во-вторых, мизерные потоки ионов через мембрану, потирбно для установления потенциала, не нарушают электронейтральности цитоплазмы и внеклеточной жидкости в целом, распределение зарядов происходит только в области, непосредственно прилегающей к плазматической мембраны.
Уравнение Нернста
Равновесный потенциал для определенного иона, например для калия, можно рассчитать по уравнению Нернста, что выглядит так:
,где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, o, i — концентрация калия снаружи и внутри клетки соответственно. Поскольку описанные процессы происходят при температуре тела — 310 ° К, а десятичными логарифмами в исчислении пользоваться легче чем натуральными, это уравнение превращают следующим образом:
Подставляя концентрации К + в уравнение Нернста получаем равновесный потенциал для калия, составляет -90 мВ. Поскольку по нулевой потенциал принимается внешняя сторона мембраны, то знак минус означает, что в условиях равновесного калиевого потенциала внутренняя Сторн мембраны сравнительно более электроотрицательным. Аналогичные расчеты можно провести и для равновесного Натиева потенциала, он составляет 62 мВ.
Уравнения Голдмана
Хотя равновесный потенциал для ионов калия составляет -90 мВ, МПС нейрона несколько менее отрицательный. Эта разница отражает незначительное но постоянное следование ионов Na + через мембрану в состоянии покоя. Поскольку концентрационный градиент для натрия противоположный такого для калия, Na + движется внутрь клетки и сдвигает суммарный заряд на внутренней стороне мембраны в положительную сторону. На самом деле МПС нейрона составляет от -60 до -80 мВ. Это значение значительно ближе к Е K чем до Е Na, потому что в состоянии покоя в нейроне открыто много калиевых каналов и очень мало натриевых. Также на встанвлення МПС влияет движение ионов хлора. В 1943 году Дэвид Голдаман предложил усовершенствовать уравнение Нернста так, чтобы оно отражало влияние различных ионов на мембарнний потенциал, в этом уравнении учитывается относительная проницаемость мембраны для каждого типа ионов:
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, [ион] o, [ион] i — концентрации ионов внутри и внутри клеток, Р — относительная проницаемость мембраны для соответствующего иона. Значение заряда в данном уравнении не сохраняется, но оно учтено — для хлора внешняя и внутренняя концентрация поменяны местами, так как его заряд 1.
Значение мембранного потенциала покоя для различных тканей
- Разделенными мышцы -95 мВ;
- Непосмугованих мышцы -50 мВ;
- Астроглией от -80 до -90 мВ;
- Нейроны -70 мВ.
Роль натрий-калиевого насоса в формировании МПС
Мембранный потенциал покоя может существовать только при условии неравномерного распределения ионов, обеспечивается функционированием натрий-калиевого насоса. Кроме того, этот белок делает также электрогенных властовости — он переносит 3 катионы натрия в обмен на 2 ионы калия, движущихся внутрь клетки. Таким образом, Na + -K + -АТФазы снижает МПС на 5-10 мВ. Подавление деятельности этого белка приводит к незначительному (на 5-10 мВ) мгновенного повышения мембранного потенциала, после чего он еще некоторое время будет существовать на достаточно стабильном уровне, пока будут сохраняться градиенты концентраций Na + и K +. Впоследствии эти градиенты начнут уменьшаться, вследствие проникнсоти мембраны к ионам, и через несколько десятков минут электрический потенциал на мембране исчезнет.
«Мембранный потенциал»
Выполнила Четверикова Р
Студентка 1 курса
биолого-почвенного факультета
Введение
Немного истории
Электричество в клетке
Мембранный потенциал
Потенциал действия
Порог раздражения
Характерные свойства потенциала действия
Заключение
Введение
Современная наука развивается стремительно, и чем больше мы движемся по пути прогресса, тем больше убеждаемся в том, что для решения каких-либо научных задач необходимо объединять усилия и достижения сразу нескольких отраслей науки.
Ранее господствовала концепция витализма, согласно которым биологические явления принципиально непостижимы на основе физики и химии, так как существует некая «жизненная сила», или энтелехия, не подлежащие физическому истолкованию. В 20 веке великий физик Бор рассматривал проблему взаимоотношения биологии и физики на основе концепции дополнительности, частным случаем которой является принцип неопределенности квантовой механики.
Бор считал, что ни один результат биологического исследования не может быть однозначно описан иначе как на основе понятий физики и химии. Развитие молекулярной биологии привело к атомистическому истолкованию основных явлений жизни - таких как наследственность и изменчивость. В последние десятилетия успешно развивается и физическая теория целостных биологических систем, основанная на идеях синергетики. Эрвин Шредингер пришел к оптимистическому, хотя и не вполне успокоительному выводу: «Хотя современные физика и химия не могут объяснить процессы, происходящие в живом организме, нет никаких оснований сомневаться в возможности их научного объяснения». Сегодня имеются все основания утверждать, что современная физика не встречается с границами своей применимости к рассмотрению биологических явлений. Трудно думать, что такие границы обнаружатся в будущем.
Напротив, развитие биофизики как части современной физики свидетельствует о ее неограниченных возможностях.
На этом примере можно наглядно проследить, как
достижения физики помогли ученым понять такое сложное явление.
Немного истории
Электричество у живых организмов человек обнаружил еще в глубокой древности. Вернее сказать, почувствовал, не подозревая при этом о его существовании. Этого понятия тогда не существовало. Например, древние греки остерегались встречаться в воде с рыбой, которая, как писал великий ученый Аристотель, "заставляет цепенеть животных". Рыба, наводившая страх на людей, была электрическим скатом и носила имя "торпедо". И только двести лет назад ученые поняли наконец природу этого явления.
Ученые давно хотели понять, какова же природа сигналов, перетекающих по нервам. Среди множества теорий возникавших в середине XVIII века, под влиянием всеобщей увлеченности электричеством, появилась теория о том, что по нервам передается ""электрический флюид"".
Идея летала в воздухе. Луиджи Гальвани, изучая грозовые разряды, использовал нервно-мышечный препарат лягушки. Подвесив его на медном крючке на ограждении балкона, Гальвани заметил, что когда лапки лягушки касались железного ограждения, происходило мышечное сокращение. На основании этого Гальвани делает вывод, что в биологическом объекте существует электрический сигнал. Однако, современник Гальвани - Алессандро Вольта исключил биологический объект и показал, что электрический ток может быть получен при контакте набора металлов, разделенных электролитом(вольтов столб). Так был открыт химический источник тока(названный, однако, позже, в честь его научного противника гальваническим элементом).
Этот спор был началом электробиологии. И вот через полвека немецкий физиолог Э. Дюбуа-Реймон подтвердил открытие Гальвани, продемонстрировав наличие электрических полей в нервах с помощью усовершенствованной электроизмерительной аппаратуры. Ответ на вопрос, как появляется электричество в клетке, был найден еще через полвека.
Электричество в клетке
В 1890 году Вильгельм Оствальд, занимавшийся
полупроницаемыми искусственными пленками предположил, что полупроницаемость
может быть причиной не только осмоса, но и электрических явлений. Осмос
возникает тогда, когда мембрана избирательно проницаема, т.е. пропускает одни
частицы и не пропускает другие. Чаще всего проницаемось мембраны зависит от размера
частицы. Такими частицами могут быть и ионы. Тогда мембрана будет пропускать
ионы только одного знака, например, положительного. Действительно, если
посмотреть на формулу Нернста для диффузионного потенциала Vд
возникающего на границе двух растворов с концентрациями электролита С1 и С2:
где u - скорость
более быстрого иона, v - скорость более медленного иона, R -
универсальная газовая постоянная, F - число
Фарадея, T -
температура, и предположить, что мембрана для анионов не проницаема, то есть v = 0, то
можно видеть, что должны появляться большие значения для Vд
![]() (2)
(2)
Потенциал на мембране, разделяющей два раствора
Таким образом, Оствальд объединил формулу Нернста и знание о полупроницаемых мембранах. Он предположил, что свойствами такой мембраны объясняются потенциалы мышц и нервов и действие электрических органов рыб.
Мембранный потенциал (потенциал покоя)
Под мембранным потенциалом понимают
разность потенциалов между внутренней (цитоплазматической) и наружной
поверхностями мембраны
![]()
С помощью электрофизиологических исследований было доказано, что в состоянии физиологического покоя, на наружной поверхности мембраны имеется положительный заряд, а на внутренней поверхности - отрицательный.
Юлиус Бернштейн создал теорию, согласно которой разноимённость зарядов определяется различной концентрацией ионов натрия, калия, хлора внутри и за пределами клетки. Внутри клетки в 30-50 раз выше концентрация ионов калия, в 8-10 раз ниже концентрация ионов натрия и в 50 раз меньше ионов хлора. Согласно законам физики, если бы живая система не регулировалась, то концентрация этих ионов сравнялась бы с обеих сторон мембраны и мембранный потенциал бы исчезал. Но этого не происходит, т.к. мембрана клетки - это активная транспортная система. В мембране имеются специальные каналы для того или иного иона, каждый канал специфичен и транспорт ионов внутри и за пределы клетки является в значительной мере активным. В состоянии относительного физиологического покоя натриевые каналы закрыты, а калиевые и хлорные - открыты. Это приводит к тому, что калий выходит из клетки, а хлор заходит в клетку, в результате этого увеличивается количество положительных зарядов на поверхности клетки и уменьшается количество зарядов внутри клетки. Таким образом, на поверхности клетки сохраняется положительный заряд, а внутри - отрицательный. Такое распределение электронных зарядов обеспечивает сохранение мембранного потенциала.
молекулярный биология мембрана потенциал
Потенциал действия

Это приводит к тому, что на внутренней поверхности мембраны накапливаются положительные заряды, а на наружной - отрицательные заряды. Такое перераспределение зарядов называется деполяризацией.
В этом состоянии клеточная мембрана существует
недолго (0,1-5 м.с.). Для того, чтобы клетка опять стала способной к
возбуждению, её мембрана должна реполяризироваться, т.е. вернуться в состояние
потенциала покоя. Для возвращения клетки к мембранному потенциалу, необходимо
«откачать» катионы натрия и калия против градиента концентрации. Такую работу
выполняет натриево-каливый насос, который восстанавливает исходное состояние
концентрации катионов натрия и калия, т.е. восстанавливается мембранный
потенциал.
Порог раздражения
Для возникновения деполяризации и последующего возбуждения раздражитель должен иметь определённую величину. Минимальная сила действующего раздражителя, способного вызвать возбуждение, называется порогом раздражения. Величина выше пороговой называется сверхпороговой, а ниже пороговой - подпороговой. Возбудимые образования подчиняются закону «всё или ничего», это значит, что при нанесении раздражения по силе, равной пороговой, возникает максимальное возбуждение. Раздражение ниже подпороговой силы не вызывает раздражение.
Для характеристики силы действующего раздражителя от времени его действия, выводят кривую, которая отражает, сколько времени должен действовать пороговый или сверхпороговый раздражитель, чтобы вызвать возбуждение. Действие раздражителя пороговой силы вызовет возбуждение только в том случае, если данный раздражитель будет действовать определенное время. Минимальная сила тока или возбуждения, которые должны действовать на возбудимые образования, чтобы вызвать раздражение называется реобазой. Минимальное время, в течении которого должен действовать раздражитель силой одной реобазы, чтобы вызвать возбуждение называется минимальным полезным временем.
Величина порога раздражения зависит не только от длительности действующего стимула, но и от крутизны нарастания. При уменьшении крутизны нарастания раздражителя ниже определённой величины, возбуждения не возникает, до какой бы силы мы не довели раздражитель. Это происходит потому, что в месте нанесения раздражителя постоянно повышается порог, и до какой бы величины не довели раздражитель, возбуждения не возникает. Такое явление-приспособление возбудимого образования к медленно нарастающей силе раздражителя называется аккомодацией.
Разные возбудимые образования имеют разную скорость аккомодации, поэтому чем выше скорость аккомодации, тем крутизна нарастания раздражителя выше.
Этот же закон работает не только для
электростимуляторов, но и для других (химических, механических
раздражителей/стимуляторов).
Характерные свойства потенциала действия
Полярный закон раздражения.
Это закон впервые был открыт П.Ф. Флюгером. Он установил, что постоянный ток обладает полярным действием на возбудимую ткань. Это выражается в том, что в момент замыкания цепи, возбуждение возникает только под катодом, а в момент размыкания - под анодом. Причем под анодом, при размыкании цепи, возбуждение значительно выше, чем при замыкании под катодом. Это обусловлено тем, что положительно заряженный электрод (анод) вызывает гиперполяризацию мембраны, когда поверхности касаются катода(отрицательно заряженного), он вызывает деполяризацию.
Закон «всё, или ничего»
Согласно этому закону, раздражитель подпороговой силы не вызывает возбуждения (ничего); при пороговом раздражении, возбуждение принимает максимальную величину (всё). Дальнейшёё увеличение силы раздражителя не усиливает возбуждения.
Долгое время полагали, что этот закон является общим принципом возбудимой ткани. При этом считали, что «ничего» - это полное отсутствие возбуждения, а «всё», - это полное проявление возбудимого образования, т.е. его способность к возбуждению.
Однако, с помощью микроэлектронных исследований было доказано, что даже при действии подпорогового раздражителя в возбудимом образовании происходит перераспределения ионов между наружной и внутренней поверхностями мембраны. Если с помощью фармакологического препарата повысить проницаемость мембраны для ионов натрия или снизить проницаемость для ионов калия, то амплитуда потенциалов действия повышается. Таким образом, можно заключить, что этот закон должен рассматриваться лишь, как правило, характеризующее особенности возбудимого образования.
Проведение возбуждения. Возбудимость.
В демиелинизированных и миелинезированных волокнах возбуждение передается по-разному, это обусловлено анатомическими особенностями данных волокон. Миелинизированные нервные волокна имеют перехваты Ранвье. Передача сигналов через такие волокна осуществляется с помощью перехватов Ранвье. Сигнал проскакивает через миелинизированные участки, и тем самым, проведение возбуждения по ним происходит быстрее, чем в немиелинизированных участках, возврат импульса обратно невозможен, поскольку в предыдущем перехвате повышается порог раздражений.
Возбудимость - это способность такни на раздражение или возбуждение и, следовательно, возникновением потенциала действия. Чем порог раздражения выше, тем возбуждение выше, и наоборот.
Величина порога раздражения находится в обратной зависимости от длительности (t) действия стимула и крутизны нарастания его силы
Таким образом, мы видим, что без помощи физики не удалось бы открыть тайну электричества в живых организмах, передачу нервных импульсов, мембранный потенциал - одни из важнейших аспектов современной биологии.